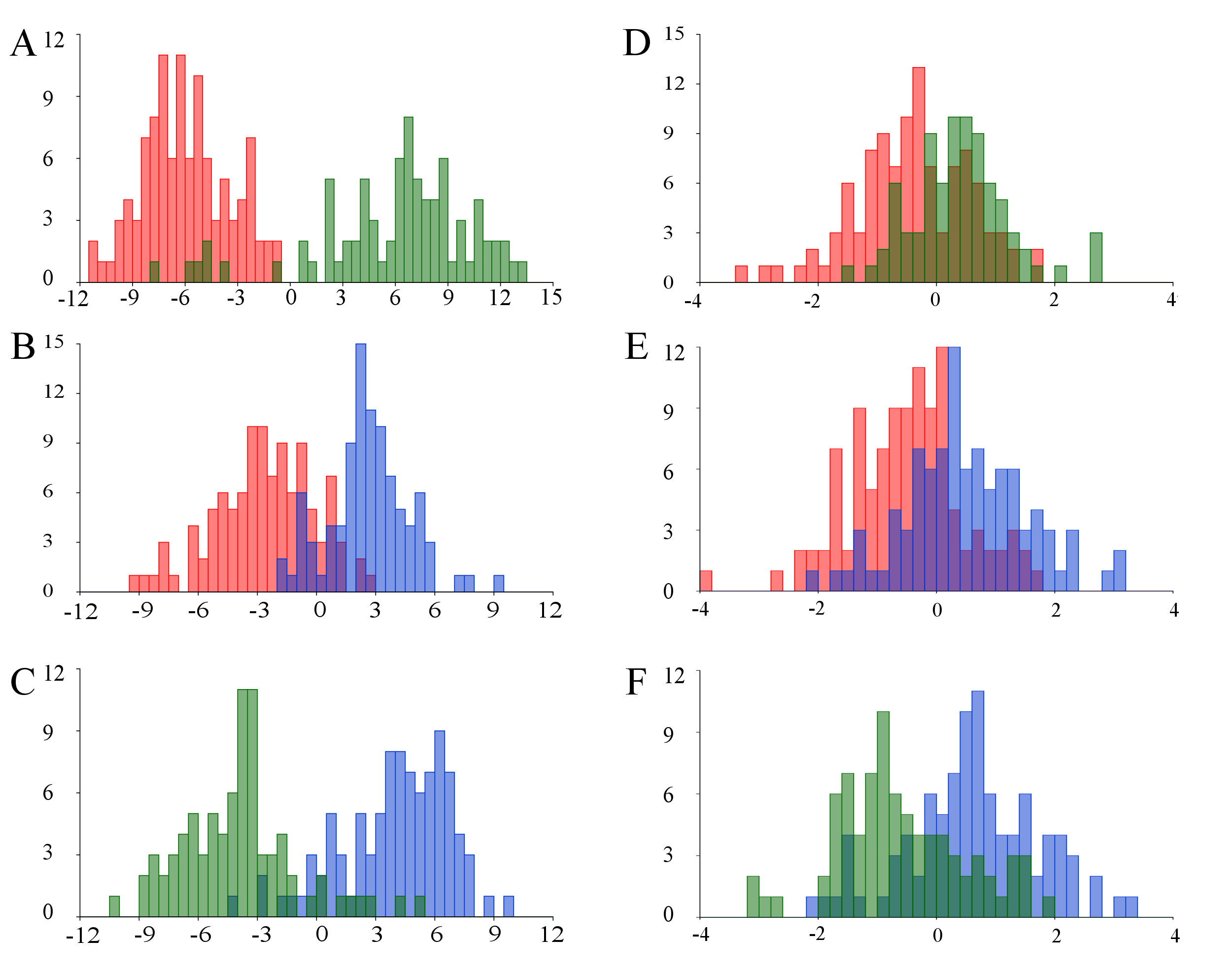
INTRODUCTION _Carollia_ is a genus of fruit-eating phyllostomid bats with a widespread distribution and high abundance from the Neotropic. These species are crucial for different ecological processes, like seed dispersal and restoration of tropical forests. Due to their ecological plasticity, species of this genus are often found in disturbed and transformed ecosystems that are under high levels of pressure. It is also common two find two or even three sympatric species all along South America. Currently, up to eight species are accepted in this genus (_C. benkeithi_, _C. brevicauda_, _C. castanea_, _C. manu_, _C. monohernandezi_, _C. perspicillata_, _C. subrufa_, _C. sowelli_), and it has been discussed the taxonomic identity of another one (_C. colombiana_). The taxonomy of these species is unclear based on morphological and morphometric data, and some of them have been considered to be complex of species yet unsolved. This genus has been of particular interest in Colombia -highest phyllostomid species richness in the world-, where two species (_C. colombiana_ and _C. monohernandezi_) have been described in the last 15 years, and at least four of the six South American species of this genus are present. A broad variety of studies have focused in this genus, offering detailed information regarding their evolutionary history, behavior, functional morphology and ecology. Also, other studies have tried to shed light on its taxonomy, studying the morphological boundaries between species, as well as the effect of both biotic and environmental traits on its morphological plasticity. Despite the lack of concise agreement, the general consensus sets size variations as the main source of morphological differentiation between species. Hence, this feature has been widely used as a taxonomic clue to identify some of these species. However, published research using molecular approaches indicate that the morphological description and discrimination of these species is inaccurate, and misleads researchers from solving the identity of unknown cryptic species. Given the fact that is becoming more evident that the use of Traditional Morphometrics (TM) for taxonomic purposes is not as accurate as thought before, the implementation of Geometric Morphometrics (GM) in this field is increasing. GM enables the decomposition of morphological variation in two independent components: size and shape. Although it has been accepted that size is the main source of variation in this genus, recent studies have re-evaluated this assumption, finding enough evidence to argue this hypothesis and propose that shape is also important for species delimitation in _Carollia_. Within GM and the variety of tools and approaches available to study morphological variation, analyzing the presence and patterns of asymmetry on a feature is been used to study hidden sources of variation (e.g. developmental instability, genetics and phenotypic variation). There are three main types of asymmetry; Fluctuating Asymmetry (FA), Directional Asymmetry (DA) and Antisymmetry (AS). Each type has specific mathematical basis and ecological interpretation: FA reflects genetic and developmental factors and it is usually used as an indicator of stress or environmental instability; DA serves to describe specific patterns of morphological variation within a sample, and it is usually associated with the inherent biology of each taxon. The utility of decomposing the morphological variation in its symmetric and asymmetric components in order to elucidate differences between species has not been sufficiently explored. Previous studies show the adequacy of this approach to detect specific patterns of morphological variation that could even be useful to taxonomically discriminate species within a genus. This methodology has only been used to study the morphology of taxa with fractal and radial symmetry. In order to further explore the importance of shape variation for the morphological delimitation and identification of _Carollia_ species, in this study, it was aimed to isolate and study the patterns of symmetric and asymmetric shape variation of the genus _Carollia_ in Colombia. MATERIALS AND METHODS Crania of 286 specimens (_C. brevicauda_=108; _C. castanea_=82; _C. perspicillata_=96) were photographed with a Nikon D5100 camera and a AF-S DX Nikkor 18-55mm f/3.5-5.6G VR lens in occlusal view. _C. monohernandezi_ was not considered for these analyses due to the small sample size available (known only for its holotype). Blaker (1976) methodology for estimating optimal focal distance was used to standardize the photographs and ensure their best quality. 16 paired landmarks and semilandmarks were used to describe cranial shape, following Klingenberg (2015) guidelines to study structures with object symmetry. Object symmetry divides an unique structure in two subregions based on a plane of symmetry. Two extra single landmarks were used to set the symmetry plane on the sagittal axis of the cranium. Landmark digitalization was performed using TPSDIG version 2.16. After the Procrustes coordinates dataset was obtained for the original configurations, a second dataset for the reflected mirror images of all original configurations was obtained in order to perform the Generalized Procrustes Analysis (GPA). GPA is a statistical method to superimpose landmarks coordinates, while removing the influence non-morphological sources of variation as a result of scaling, rotation and translation. A consensus, tangent configuration is obtained from the average coordinates of the original-mirror dataset. To peform a GPA in configurations with object symmetry, each original configuration has to be combined with its mirror image, after relabeling landmarks of the mirror configuration, to ensure the correct pairing between original and mirror landmarks. Further, a Procrustes ANOVA was performed in order to test statistical differences between species, as well as statistical presence of DA. Statistical significance on the “Side” component of the Procrustes ANOVA means that there is a consistent, asymmetrical distribution in the variation between both sides of the cranium, which indicates the presence of DA. Using the Procrustes ANOVA test makes the assumption that data points have a multivariate normal distribution in the tangent space, and that the variation is isotropic (equally distributed in all directions of the variation). This assumption has been considered to be inaccurate when studying biological structures. Hence, using alternative analyses, like MANOVA is recommended. MANOVA was performed to check the presence of FA and DA without the assumption of isotropic variation. When interpreting the MANOVA, statistical significance for the “Ind*Side” factor means that the morphological variation is due to the interaction between the within-group variation and the variation on each side of the cranium, reflecting presence of FA. Procrustes ANOVA and MANOVA were conducted using the MorphoJ software. A Canonical Variate Analysis (CVA) was used to assess the differences between the potential of the asymmetric and symmetric components to discriminate all three species. CVA is a multiple discriminant function that groups individuals within a sample into pre-established groups, based on multivariate data. CVA was performed for the symmetric and asymmetric component separately. All CVA analysis were conducted using MorphoJ. Finally, using the Procrustes coordinates, thin-plate splines were obtained to illustrate more effectively the morphological differences in the asymmetric and symmetric variation between groups. This was repeated for each species, and each Canonical Variate of each CVA. RESULTS Procrustes ANOVA provided statistical evidence of significant differences between species, and confirmed the presence of DA in the sample (Table 1). The “Sp*Side” factor was not significant, but since the mathematical interpretation of this factor does not reflect the presence of any kind of asymmetry or any other biologically relevant feature, this factor was excluded of further interpretations. MANOVA showed statistical significance for the presence of both DA and FA, using a non-isotropic model of variation. Because this approach avoids the isotropic variation model, considers only the variation in the shape of the structure, making it statistically a more powerful tool. By comparing the graphical output of the CVAs obtained for the symmetric and asymmetric component of the variation, it was clear that the asymmetric component is more accurate to discriminate the three species. The symmetric component was not useful to discriminate neither of the three species, overlapping almost completely the species all along the morphospace. Despite the presence of a certain degree of overlapping between species in the asymmetric component, most of the samples were accurately discriminated, suggesting that morphological differences between these species are better represented in their asymmetric variation. The overlapping found on both components opens the question of what other mechanisms and patterns rely behind the morphological variation on _Carollia_ species. Thin-plate splines for the canonical variates also showed differences on the distribution of both components of the variation along the cranium. While both canonical variates of the asymmetric component varied greatly in the Neurocranium, canonical variates of the symmetric component showed a slight variation in the Splachnocranium. Species-specific patterns of asymmetric morphological variation were evident after comparing the thin-plate splines obtained for the species. Asymmetric variation on all three species was predominantly concentrated in the Neurocranium, specifically in the occipitomastoid suture region. Variation was greater at the anterior region of the occipitomastoid suture for _C. castanea_, at the middle section for _C. brevicauda_, and at the posterior region for _C. perspicillata_. DISCUSSION With this study, morphological variation in bilateral organisms was decomposed in its asymmetric and symmetric components for the first time, using principles of the methodology designed to study organisms with complex symmetries. Our results support other recent studies that discuss the importance of shape variation for the discrimination of species and reject the assumption that size is the best way to distinguish _Carollia_ species. This also suggests that the complex morphological delimitation and boundaries of _Carollia_ species could be better understood by analyzing its asymmetric component, rather than focusing in traditional approaches that are restricted to symmetric variation. In this study, presence of FA was interpreted as a signal of high phenotypical diversity within species, rather than an indirect indicator of ecosystem variability, DI or some kind of stress (e.g. genetic or environmental). In the asymmetric CVA, specimens of _C. brevicauda_ and _C. perspicillata_ dispersed similarly across the morphospace, while specimens of _C. castanea_ dispersed slightly less. Also, species-specific patterns of morphological variation found in the thin-plate splines revealed some similarities between _C. brevicauda_ and _C. perspicillata_, as well as _C. castanea_ as the most morphologically distant species.