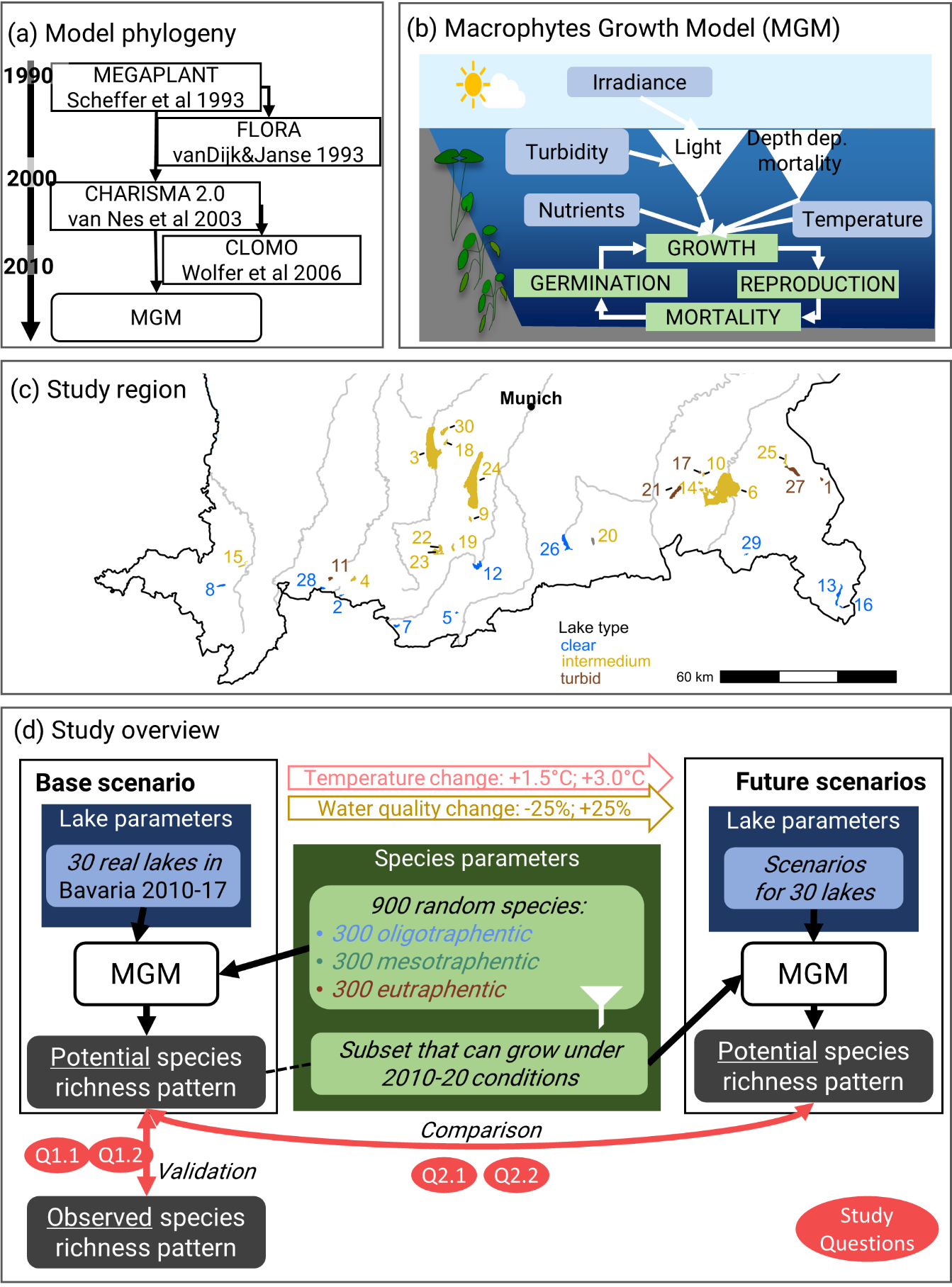
IntroductionSubmerged macrophytes provide a broad range of ecosystem services in lakes (Thomaz, 2021). They create habitat for many other species and change the lake environment by binding nutrients or stabilising the sediment. Most importantly, the eco-physiological processes controlling their growth and survival are strongly affected by environmental conditions, as they depend on light availability, nutrient availability, and temperature. However, the accelerating global biodiversity loss, especially of submerged macrophytes, is well documented (Körner, 2002; Phillips et al., 2016; Sand‐Jensen et al., 2000; Zhang et al., 2017) despite evidence of increasing species richness in some lakes (Murphy et al., 2018). The main influencing factors on the change of species richness seem to be global climate and regional land-use changes (Hofstra et al., 2020; Zhang et al., 2017) resulting in changes of light availability (due to changes in water turbidity), nutrient availability, or water temperature. The ways in which climate change influences water temperature, nutrients, and turbidity in lakes are highly complex including direct and indirect effects (Lind et al., 2022). For example, lake water temperature rises due to climate change which can also have indirect impacts on turbidity and nutrients in lakes, as higher temperature can promote algae growth (lower light availability). Therefore, nutrients and turbidity seem to co-vary, while temperature could change independently. However, nutrient content and turbidity can also be influenced by land-use practices (e.g. content and timing of fertiliser on fields), and wastewater treatment management. Overall, the direction and impact of future climate change on submerged macrophytes seems to be more obvious than changes in land use.All these stressors have an effect on the eco-physiological processes of macrophytes (Cao & Ruan, 2015; Reitsema et al., 2018). Hence, changes in these stressors likely affect the geographical distribution of individual species and of species richness of submerged macrophytes. Because of their response to their physio-chemical, geomorphological, hydrological, and biotic surroundings (O’Hare et al., 2018), species can be differentiated into oligo-, meso-, and eutraphentic species by their preference for nutrient conditions (Melzer, 1999). The presence of distinct species can then be an indicator of the water quality and ecological state of the lake (Schaumburg et al., 2004). However, studies on macrophyte species richness distribution remain largely based on observational and correlative studies, while there is a need to understand how simultaneous stressors combine and may result in synergistic effects on biodiversity (Lind et al., 2022). Therefore, it is paramount to assess these potential effects on macrophytes.Process-based models based on first principles and known ecophysiology are better suited to assess biodiversity response from changing conditions than correlative models (Cabral et al., 2017; Dormann et al., 2012; Higgins et al., 2020; Schouten et al., 2020). To predict the potential distribution of species based on environmental factors, process-based models incorporating critical eco-physiological processes are necessary. The application of process-based models describing the growth of submerged macrophytes have a long tradition, already stemming as early as the late 1980s (Best et al., 2001; Collins & Wlosinski, 1989; Herb & Stefan, 2003; Hootsmans, 1994; Scheffer et al., 1993; Wortelboer, 1990). The majority of those models were developed to answer different study questions, however, like the effect of macrophytes on algal blooms (Asaeda & Van Bon, 1997), the effect of varying light regimes (Herb & Stefan, 2003), or their impact on water quality (Sachse et al., 2014). Furthermore, most models were only used and calibrated for one or a few species (Gao et al., 2017; van Nes et al., 2003) and under very specific environmental conditions. None of these models was used to study the macroecological distribution patterns of macrophytes or its response to environmental change. Among the reasons for the delay in applying any of these models to multiple species and under different environmental conditions is the lack of computationally efficient models and empirical data to constrain both eco-physiological and environmental parameters. Hence, applying eco-physiological models to assess the species and richness distribution in both current and future conditions deserves further attention for this neglected group of species.Macrophytes are still underrepresented in trait-based research and in environmental change assessments (Dalla Vecchia et al., 2020; Iversen et al., 2022). Consequently, the determination of a broad range of eco-physiological parameters has yet to be established for most macrophytes. In the case of low trait-based information, applying eco-physiological models to a virtual species pool remains the best alternative to assess impacts of environmental change on macrophytes (Cabral et al., 2017; Dormann et al., 2012). As computational power and methods are evolving (Peréz-Sánchez et al., 2015), experiments with a broad range of randomly selected species within defined functional types can be a way of determining trait combinations (potential species) that allow species to survive and reproduce, as already done for terrestrial plants (Webb et al., 2010; Zakharova et al., 2019). In such applications, the process-based model acts as a performance filter, with the surviving virtual species representing those trait combinations able to cope with the environmental conditions given the considered mechanisms. However, this approach was not yet used for macrophytes.In this study, we tackle two main objectives. First, we address the potential species richness of oligotraphentic, mesotraphentic, and eutraphentic submerged macrophytes under recent environmental conditions. We ask: How many observed and potential species of the species pool can grow in clear, intermediate, and turbid lakes (Q1.1)? Do the potential species richness patterns across the depth follow the observed distribution in all lake types (Q1.2)? Second, we assess scenarios of water temperature increase and water quality change (increase or decrease in both nutrients and turbidity). Here, we ask: In which depth and lake types do we lose or gain oligo-, meso-, and eutraphentic species (Q2.1)? Is this change dependent on eco-physiological traits (Q2.2)?To answer questions Q1.1 and Q1.2, we run random species parameter combinations within the three defined parameter spaces of oligotraphentic, mesotraphentic, and eutraphentic species and analyse the resulting distribution patterns of the growing species by comparing them with the corresponding observed pattern. We expect to find the highest species richness in moderately nutrient rich lakes (Q1.1) (Lewerentz & Cabral, 2021). We hypothesise to find hump-shaped patterns (Q1.2) of species richness along depths (Lewerentz et al., 2021). In general, we hypothesise that we see a higher potential species richness than observed species richness because limiting processes like herbivory have not been modelled (Q1.1 and Q1.2). To answer question Q2.1 and Q2.2, we run scenarios of water temperature increase and scenarios of water quality change for the recent potential species pool. We hypothesise that the studied lakes will lose species under increased turbidity and nutrient conditions but gain species under decreased turbidity and nutrient conditions and under increased water temperature (Q2.1) (Lewerentz & Cabral, 2021). To answer question Q2.2, we determine the plant traits that significantly influence if a species will win or lose habitat within two selected scenarios of turbidity and nutrient decrease or increase. We hypothesise that under increased conditions, high biomass production is the main advantage, as species can grow at high rates even under limited conditions.