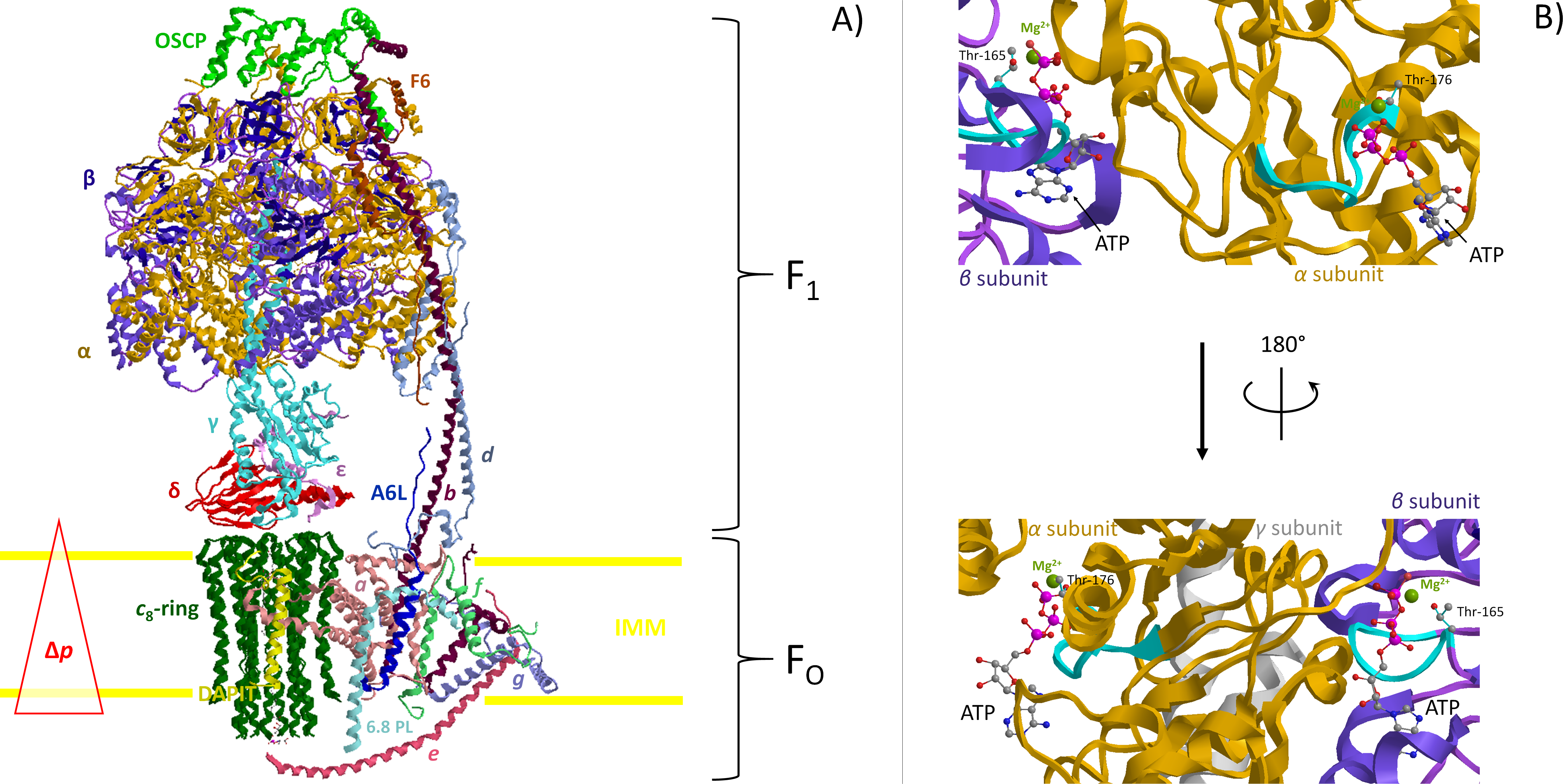
The mitochondrial F1FO-ATPase in the presence of the natural cofactor Mg2+ acts as the enzyme of life by synthesizing ATP, but it can also hydrolyze ATP to pump H+. Interestingly, Mg2+ can be replaced by Ca2+, but only to sustain ATP hydrolysis and not ATP synthesis. When Ca2+ inserts in F1, the torque generation built by the chemomechanical coupling between F1 and the rotating central stalk was reported as unable to drive the transmembrane H+ flux within FO. However, the failed H+ translocation is not consistent with the oligomycin-sensitivity of the Ca2+-dependent F1FO-ATP(hydrol)ase. New enzyme roles in mitochondrial energy transduction are suggested by recent advances. Accordingly, the structural F1FO-ATPase distortion driven by ATP hydrolysis sustained by Ca2+ is consistent with the permeability transition pore signal propagation pathway. The Ca2+-activated F1FO-ATPase, by forming the pore, may contribute to dissipate the transmembrane H+ gradient created by the same enzyme complex.