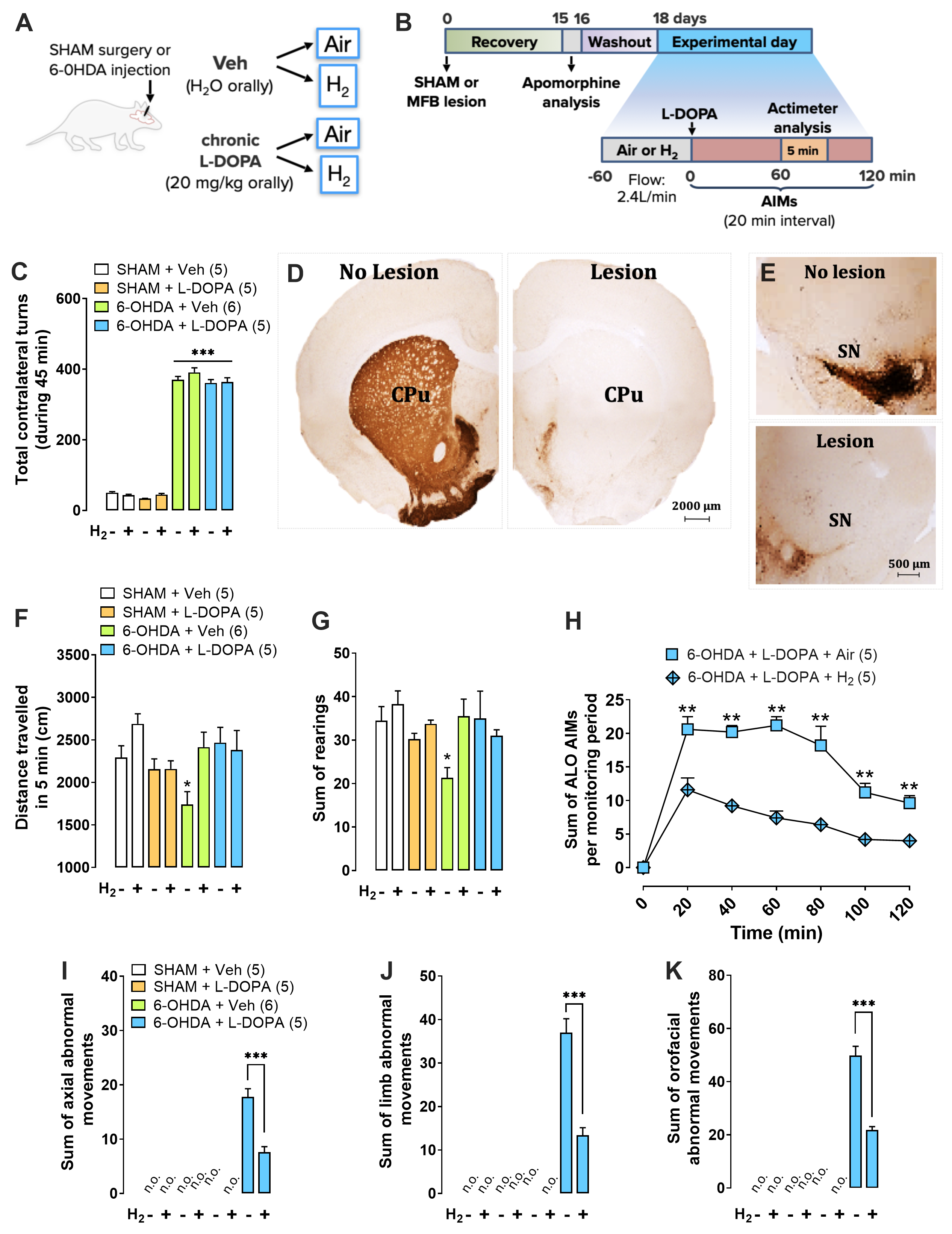
Bruna M. Santos PhD1,2; Glauce C. Nascimento PhD1,3; João F. Pedrazzi PhD 4; Danyelle Silva-Amaral MS1 ; Mariza Bortolanza PhD3; Grant T. Harris BS2; Luiz G. S. Branco PhD 1,3*, Elaine Del Bel PhD1,3,4*1 Department of Physiology, Medical School of Ribeirão Preto, University of São Paulo, Ribeirão Preto, SP, Brazil.2Thermoregulation and Systemic Inflammation Laboratory (FeverLab), St. Josephs’ Hospital and Medical Center, Phoenix, AZ, USA.3 Department of Basic and Oral Biology, Ribeirão Preto Dentistry Faculty, University of São Paulo, Ribeirão Preto, SP, Brazil.4 Neuroscience Graduate Program, Medical School of Ribeirão Preto, University of São Paulo, Ribeirão Preto, SP, Brazil.*Corresponding Authors :Elaine Del Bel ; email:[email protected]; Department of Basic and Oral Biology, Dental School, University of São Paulo (USP), Ribeirão Preto, SP, 14040-904, Brazil.Luiz GS Branco ; email: [email protected] ; Department of Basic and Oral Biology, Dental School, University of São Paulo (USP), Ribeirão Preto, SP, 14040-904, Brazil.Text (3692 words)- (Up to 3700 words excluding of abstract, legends, and references)Running title: The anti-inflammatory effect of H2 in LIDKey words: 6-Hydroxydopamine, Parkinson’s disease, neuroinflammation; striatum; systemic inflammationThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.Funding sources: Grant GCN [#88882.317597/2019-014, National Council for Scientific and Technological Development (PNPD-CNPq), Brazil]. Grant JFP [#88887.341956/2019-00, Programa Nacional de Pós-doutorado, Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (PNPD/CAPES)]. Grant MB [#88887.334730/2019-00, Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)]. Grant LGSB [#2016/17681-9, São Paulo Research Foundation (FAPESP), Brazil. Grant EDB [#2017/243044-0, São Paulo Research Foundation (FAPESP), Brazil]; [# 88881.198836/2018/01, Coordenação de Aperfeiçoamento de Pessoal de Nível Superior and Deutscher Akademischer Austauschdienst (CAPES/DAAD – 2019-2024), Brazil and Germain], [#88887.192409/2018-01, Coordenação de Aperfeiçoamento de Pessoal de Nível Superior and Comitê Francês de Avaliação da Cooperação Universitária com o Brasil (CAPES/COFECUB- 2019-2024), Brazil and France.], [#302434/2019-04, National Council for Scientific and Technological Development (CNPq), Brazil].Abstract (226 words)Background: L-3,4-dihydroxyphenylalanine (L-DOPA)-induced dyskinesia is a side effect of Parkinson’s disease treatment and it is characterized by atypical involuntary movements. A link between neuroinflammation and L-DOPA-induced dyskinesia has been documented. Molecular hydrogen has neuroprotective effects in Parkinson’s disease models and has a major anti-inflammatory effect.Objective: To test the hypothesis that molecular hydrogen inhalation reduces L-DOPA-induced dyskinesia.Methods: 15 days after 6-hydroxydopamine lesions of dopaminergic neurons were made (microinjection into the medial forebrain bundle), chronic L-DOPA treatment (15 days) was performed. Rats were exposed to molecular hydrogen (2% gas mixture, 1h) or air (controls) before L-DOPA injection. Abnormal involuntary movements and locomotor activity were conducted. Striatal and plasma samples for molecular and morphological evaluations were collected after the abnormal involuntary movements analysis.Results: Molecular hydrogen inhalation attenuated L-DOPA-induced dyskinesia. The gas therapy did not impair the improvement of locomotor activity achieved by L-DOPA treatment. Display of abnormal involuntary movements was positively correlated with plasma IL-1β and striatal TNF-α levels and negatively correlated with striatal IL-10 levels. H2 inhalation reduced activated microglia in the lesioned striatum, which is consistent with the observed reduced pro-inflammatory cytokines levels.Conclusions and implications: Prophylactic molecular hydrogen inhalation decreases abnormal involuntary movements in a preclinical L-DOPA-induced dyskinesia model. The molecular hydrogen antidyskinetic effect was associated with decreased striatal and peripheral inflammation. This finding has a translational importance to L-DOPA-treated parkinsonian patients’ well-being.IntroductionSeveral motor and non-motor debilitating symptoms of Parkinson’s disease (PD) are generated by dopamine (DA) depletion. Levodopa (L-DOPA) is recognized as the most effective and well tolerated drug for PD treatment. However, long-term L-DOPA therapy has been associated with the development of abnormal involuntary movements referred to as L-DOPA-induced dyskinesia (LID)1. The development of dyskinesia represents a serious side effect that negatively affects quality of life of people with PD, and is also associated with both motor, including an increased risk of postural imbalance and trauma from falls, and non-motor operations, particularly, fluctuations in mood and cognitive performance2–4. Although various compounds have been used for treating LID, their success has been limited3.The pathogenesis of LID is associated with non-physiological stimulation of DA receptors using L-DOPA, which combined with the reduction of gamma-aminobutyric acid transmission pale-thalamic activity, generates neuronal hyperactivity in the striatum5–9. Moreover, recent studies suggest that astrocyte and microglia activation increase pro-inflammatory cytokines production, contributing to the appearance of LID10,11. In addition, excessive levels of glutamate and dopamine in the striatum feed this inflammatory environment, favoring LID12,13. A key unmet medical need for the PD community is a strategy for ameliorating LID10,14,15.Despite the fact that neurodegenerative disorders and associated consequences have been consistently related to central nervous system (CNS) derangement, they also have a strong associative component with environmental factors16. Given the evidence of the profound influence of the periphery producing environmental signals to the CNS, the periphery-brain axis has been increasingly explored within the neurodegenerative disorders such as Alzheimer’s and PD17,18. One of the peripheral influences that has been implicated in the neurodegenerative disorders outbreak and progression is the peripheral immune system19. Intestinal inflammation, for example, is remarkably observed in patients with PD20,21. Gut microbiota related to PD clinical phenotype produces an improper ratio of sub-products with anti-inflammatory agents such as short-chain fatty acids and molecular hydrogen (H2)22,23.Studies have shown that H2 may protect the brain from various neuronal conditions, including neurological emergencies and neurodegenerative diseases24-26. H2 in rodents protects mesencephalic dopaminergic neurons from 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MTPT)24and 6-hydroxydopamine (6-OHDA)25 induced degeneration. Several lines of evidence indicate that neuro-immune function and behavior are influenced by H226–28. Accumulating studies indicate that H2 not only has antioxidant29,30 but also anti-inflammatory properties31–33. However, there are no previous studies addressing the putative H2 benefits on LID.Hence, we tested the hypothesis that H2 inhalation reduces L-DOPA-induced dyskinesia in an animal PD model. We evaluated the effect of H2 inhalation on abnormal involuntary movements (AIMs) in PD rats treated with L-DOPA. Moreover, we evaluated the role of H2 in pro-inflammatory [tumor necrosis factor (TNF)-α, interleukin (IL)-1β, IL-6, interferon (IFN)-ϒ, and anti-inflammatory (IL-10) cytokine levels in the plasma and striatum of PD-lesioned rats treated with L-DOPA. Finally, we examined whether H2 inhalation influenced the morphology of striatal astrocytes and microglia in PD-lesioned rats chronically treated with L-DOPA.2.0. Materials and Methods More detailed description of our experimental procedures can be found in the Supplementary Methods.2.1. Animals Male Wistar rats (200-250 g, n = 65; USP-RP, SP, Brazil) were used. The experimental protocol followed the Ethical Principles in Animal Research delineated by the guidelines for the care and use of mammals in Neuroscience and Behavioral Research and was approved by the Ethics Committee on the Use of Experimental Animals of the University of São Paulo (#2016.1.667.58.4).2.2. Dopaminergic lesion with neurotoxin 6-hydroxydopamine Rats were anesthetized with 2,2,2-tribromoethanol (250 mg kg-1 ip) and fixed into the stereotaxic apparatus. The incisor bar was set at 3.3 mm below the interaural line. Rats received one injection of 2.0 μl 6-OHDA into the left medial forebrain bundle as described by Gomes et al., 20085 (6-OHDA - 2.5 µg µl-1 in 0.9% NaCl supplemented with 0.02% ascorbic acid, 1 μL min-1).Motor asymmetry was assessed 15 days after the lesion of the nigrostriatal pathway by apomorphine-induced rotational behavior analysis (0.5 mg kg in 0.9% NaCl, subcutaneous, Sigma) (Figs. 1C and 2D)34. The lesion was confirmed histologically at the end of the behavioral tests (described below) by tyrosine hydroxylase (TH) immunohistochemistry in the striatum and substantia nigra (Figs.1D and E). L-DOPA treatment started two days after the apomorphine-induced rotational test.2.3. H2 administration All animals were adapted to the chamber with ambient air at a flow rate of 2.4 L/min, one day before the experiments. On the experimental day, animals were exposed to 2% H2-mixture (21% of O2 balanced with N2) or Air-mixture (0% H2; 21% O2 N2-balanced) for 1 hour at the same flow rate as the training day. Continuous monitoring of H2 concentration in the chamber was made possible by using a gas analyzer to verify the H2 outflow from the chamber. The tested concentration of H2 in the mixture (2%) was chosen due to the consistent and repeatable anti-inflammatory effect observed in previous studies29,32,33.2.4. L-DOPA-induced abnormal involuntary movements analysisRats were subjected to chronic L-DOPA (20 mg kg plus benserazide 5mg kg, orally by gavage) administration for 15 days (once a day) to induce a stable expression of dyskinesia14. Animals showing AIMs scores greater than 10 and severity grading greater than 2 on at least one AIMs subtype during the effect of L-DOPA were selected for the behavioral study30-32. Rats were monitored individually for orofacial and limb dyskinesia, and axial dystonia or AIMs using a rat dyskinesia scale34–37. This scale considers the severity and amplitude of the axial, limb and orolingual AIMs (the scores ranged from 0 to 4) (for review see Cenci and Lundblad 200738). A dyskinesia time curve was generated by plotting global AIMs scores from each monitoring time individually for an entire testing session (180 min).2.5. Actimeter test To evaluate possible motor effects of the treatments, the spontaneous locomotor activity of each animal was individually assessed in a photoelectric actimeter (Actitrack, Panlab, Barcelona, Spain). The apparatus consists of a square arena (45 x 45 x 20 cm) equipped with 16 photocells that detect horizontal movement and 16 photocells that detect vertical movement. Thus, the locomotor activity was obtained from recording the number of interruptions of the photoelectric cells during 5 minutes. The amount of rearings was counted using total vertical activity movements.2.6. Plasma and striatum sampling The animals were euthanized by decapitation 1 hour after L-DOPA or its vehicle administration. The trunk blood was rapidly collected in heparin-coated tubes and subsequently centrifuged at 3,500 rpm for 20 min at 4°C. The brain was removed, and the rostral and medial portion of the striatum (rostral: 1.7 mm and medial: 0.7 mm from bregma) was carefully excised, frozen by submersion in dry ice chilled isopentane and The medial-caudal (medial: 0.7 mm and caudal: -0.8 mm and from bregma) portion of the striatum was collected and post-fixed in tamponade paraformaldehyde (4%) for immunohistochemistry preparation.2.7. Measurement of plasma and striatal cytokines levels The inflammatory profile of the target samples was evaluated using a high sensitivity enzyme-linked immunosorbent assay (Luminex). Luminex assay was conducted using Luminex Magpix technology (Austin, TX, USA) according to manufacturer guidelines (LXSARM - 05, R&D, Minnesota, MN, USA)39.2.8. Immunohistochemistry Serial coronal sections throughout the mediocaudal extent of the striatum (Bregma +2.76mm, Interaural 11.76mm – Bregma -2.28mm, Interaural 6.72mm) were cut (25 µm) using a freezing microtome (Leica, model CM1850). Immunohistochemistry was performed in these sections using a standard peroxidase-based method5 to quantify TH, glial fibrillary acidic protein (GFAP) and OX-42 (CD11b/c equivalent protein of microglia). The sections were incubated overnight at room temperature with the primary antibodies (Suppl. Table 1): anti-TH; anti-GFAP or anti-OX-42, followed by 2 h of incubation with biotinylated secondary antibody (1:250, Vectastain). The chromogen used was diaminobenzidine (Sigma-Aldrich, St. Louis, MO, USA) and the slices were mounted on slides and cover slipped for microscopic observations. Digital images were obtained using a Leica microscope (Leica Microsystems Launches Leica FW4000 - Cambridge, UK) under 20x (for TH optical density and OX-42/GFAP number of cells quantification) or 40x (for OX-42/GFAP morphology) objectives. The quantification of the brain area was measured using the ImageJ system (ImageJ, RRID: SCR_003070,National Institutes of Health - NIH; Schneider et al., 201240). Analysis of microglia (OX-42) and astrocyte (GFAP) immune labeling morphology was conducted as described by Giocanti-Auregan et al. (2016)41 using Fiji algorithms (RRID: SCR_002285) and a generated skeleton image. The parameters analyzed were the number of cells (per 0.1mm2), branches, the number of intersections or branching points, and the mean process length42.